The virial theorem and the Price equation
Abstract
We observe that the time-averaged continuous Price equation is identical to the positive momentum virial theorem, and we discuss the applications and implications of this connection.
The virial theorem
The virial theorem was first described by Rudolf Clausius in connection with his studies on heat transfer [6]. In its simplest form, it relates the time-averaged kinetic energy of objects with masses and velocities , to their time averaged potential energy , where are the forces acting on the objects and are their respective positions:
Theorem 1 (Virial theorem, 1870)
For stably bound gravitational systems,
| (1) |
The mathematical underpinning of the virial theorem is the product rule from calculus. The derivative of the Clausius virial where is
| (2) |
Since for stably bound systems the velocities and positions of objects have upper and lower bounds, the average of the derivative of over a period of time will be zero in the limit of large , i.e. for large , . When , we obtain from (2) the virial theorem (1): .
The virial theorem was well known to physicists in the late 19th and early 20th centuries [38, 9], however, its power as a discovery tool for astrophysics was first highlighted by Fritz Zwicky [58]. Zwicky used the virial theorem to estimate the mass of the Coma cluster, thereby identifying a mass deficit in comparison to luminosity estimates, leading him to posit the existence of what he called dunkle materie (dark matter) [58]. Although Zwicky’s mass estimates were inaccurate [48, 30], the principle of using the virial theorem to identify a measurement gap was sound, and the virial theorem has become widely used in physics and astrophysics. It can be used to derive classic laws such as the ideal gas law [14, 20], and extensions are applicable in many settings, including quantum mechanics [17], astrophysical hydrodynamics [44], and fluid mechanics [32, 1].
The Price equation
The Price equation [36] pertains to selection in evolutionary processes. It was motivated by a desire to understand the evolution of altruism [22], and has been described as a “fundamental theorem of evolution” [37] due to its generalization and unification of many results in evolutionary biology. For example, Fisher’s fundamental theorem of natural selection [13] is a special case of the Price equation [37, 15].
The Price equation relates the change in a trait in a population over time, to fitness values in subpopulations. Formally, the (discrete) Price equation as published in [36] (we follow notation from [15]) considers a numerical trait in subpopulations at time denoted . The subpopulations have sizes , and have (Wrightian) fitness defined by where denotes the time interval of one generation [52]. Let be the relative size of the population, and define the population average fitness to be . Note that forms a probability distribution for viewed as a random variable, and . Let , , and with .
Theorem 2 (The Price equation, 1970)
| (3) |
where is the expected value of the Hadamard product of and with respect to the relative subpopulation sizes, and is the covariance between the subpopulation fitnesses and trait values with respect to the relative subpopulation sizes.
Intuitively, if subpopulation fitness has positive covariance with trait values, then the trait is beneficial, and the trait value, averaged across populations, will increase after a generation. However, if the covariance between subpopulation fitness and trait values is negative, higher trait values are detrimental and the trait value averaged across populations will decrease after a generation.
The Price equation as published in [36] is discrete in time, and proof of the identity uses basic properties of expectation and covariance along with the fact that , which we leave as an exercise for the reader. Note that
The discrete-time Price equation has a continuous-time analog [35, 10]. It is formulated using the Malthusian fitness given by instead of the Wrightian fitness .
Theorem 3 (The continuous Price equation, 1972)
| (4) |
The continuous-time Price equation (4) is the continuum limit of the discrete Price equation (3). To see this, we begin by multiplying the Price equation by :
We will now see why, contrary to convention, we have indexed the variables in equation (3) with time. Starting with the left hand side, we observe that
Therefore,
The covariance term, in the limit as , is given by
Let . Note that and that . Substituting for yields
Finally, we have that
In summmary,
The Price equation from the virial theorem
In the physics setting, recall that the momentum . Let , i.e. acceleration divided by velocity. If all the momenta are greater than zero, i.e., for all , we can define relative momentum as . Consider the virial density [11], whose derivative is . The product rule applied to the virial density is
| (5) |
This shows that the virial (density) equation (5) and the continuous Price equation (4) are mathematically identical. Therefore, the relationships between traits and fitness in evolutionary biology are not only reminiscent of the relationships between physical quantities like distance, velocity, and acceleration; they are the same. It is therefore not surprising to find a direct analog of the virial theorem in genetics [16]:
Theorem 4 (Frank and Slatkin, 1990)
For a population in equilibrium where is the allelic state of a haploid genotype, for some , and ,
In other words, the rate at which selection removes phenotypic variance from the population (left-hand side) is equal to the rate at which mutation adds variance (right-hand side). Other versions of the Price equation can provide physical insight.
In summary, we have the following relationships:
| Biology | Physics | ||
| Price equation | virial theorem | ||
| continuous Price equation |
In genetics, the natural time increment to consider is discrete (generation), whereas in physics continuous-time is more natural. Thus, the discrete Price equation pertains to change in a trait after a single generation, whereas the virial theorem is formulated with continuous-time, and is additionally time averaged. However, the less intuitive forms of these equations that arise from the correspondences derived above may yield important insights. For example, the perspective of the virial theorem as a special case of the equipartition theorem [34] may be fruitful in evolutionary biology [31]. Translation between biology and physics via the virial theorem and the Price equation may also accelerate discovery of generalizations. While the stochastic Price equation in evolution [41] and the stochastic virial theorem in astronomy [7] were discovered independently, their similarity suggests other generalizations could similarly parallel each other. Moreover, the virial theorem has been applied in a variety of fields (for example economics [2]), meaning that understanding its relationship to the Price equation could be relevant beyond physics and biology.
Simple harmonic motion
In the book “Ecological Orbits: How Planets Move and Populations Grow” [18], Ginzburg and Colyvan show that a maternal effect can generate population cycles [19], obviating the need for predator-prey models to explain such dynamics. Formally, they posit that a trait that changes over time is linked to the size of a population as follows:
| (6) | |||||
| (7) |
where and are monotonically increasing functions (note that we have adjusted their notation). The maternal effect is captured by on the right-hand side of the trait evolution equation, indicating that a trait associated with individuals in a generation depends on the trait of mothers in the current generation, as well as the fraction of the total resources available to each individual in the next generation, i.e. . We extend this model to include an additional function that captures the potentially dominant impact of transgenerational effects as seen in matrotrophic species [3, 21, 39, 42]:
| (8) |
This formulation captures environmental impacts on the mother which affect her offspring directly, such as nutrition during gestation [49]. To derive a continuum limit from (8) we replace with an arbitrary time increment to obtain
| (9) |
Note that (9) reduces to (8) when . We can simplify (9) in the case where transgenerational effects dominate, i.e., we assume that and that
In this case
or equivalently
| (10) |
We note that it makes biological sense that should be a monotonically increasing concave function, since it captures the diminishing returns of increasing food per individual mother. This motivates the following specific functional form for :
| (11) |
where is a scaling factor representing the strength of the gestational maternal effect. Note that in what follows can be more general and include an additive constant, although we omit it for simplicity of presentation. Substituting (11) into (10) and defining yields
| (12) |
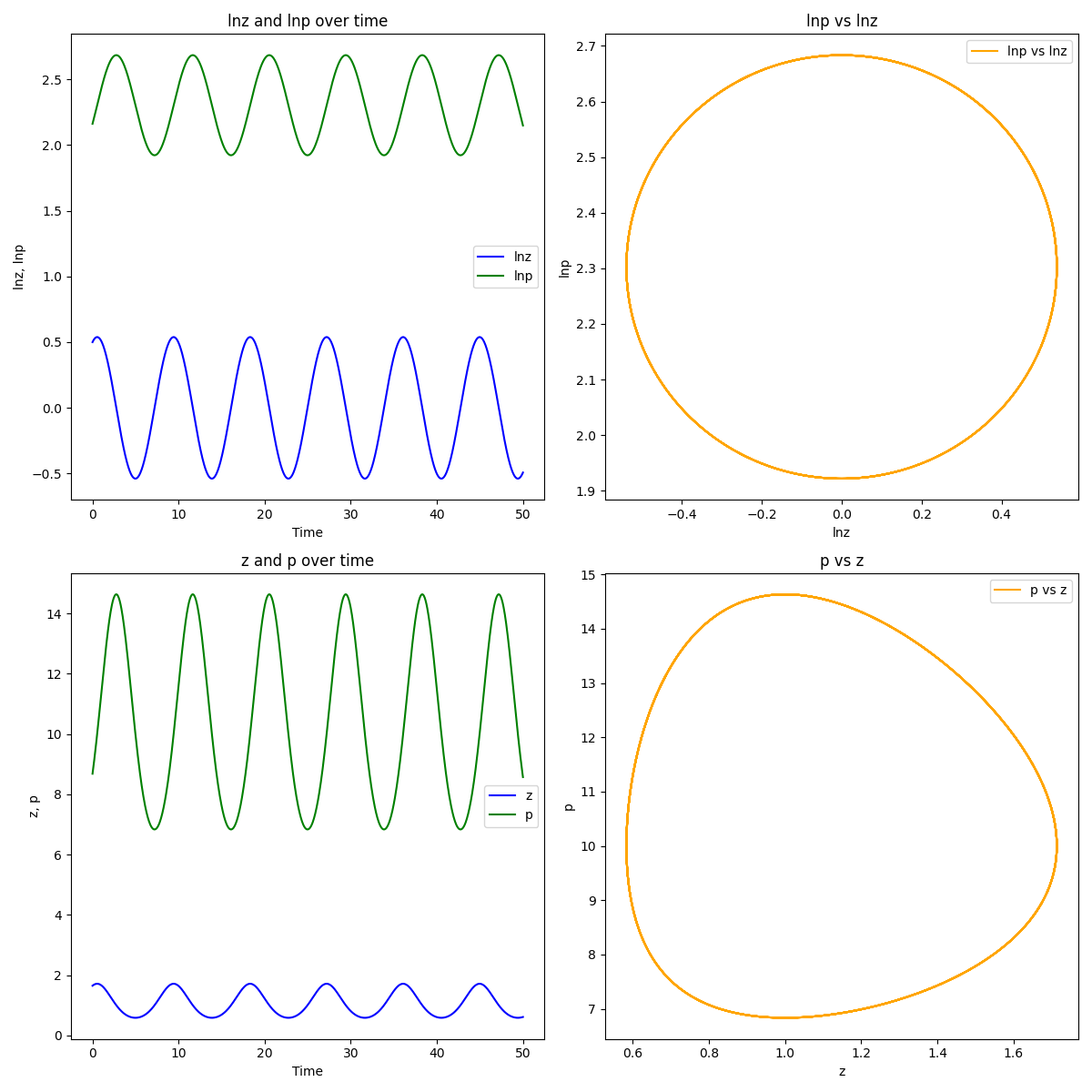
Now, as suggested by the first term in (5), we consider the relationship between Malthusian fitness and trait value . Similarly to (10), the continuum limit of (6) becomes
If we further assume that fitness is linearly related to the logarithm of the trait value, i.e. , for some constants, and , we have that:
| (13) |
Then, differentiating (12) and setting we find that
Note that this is the second order differential equation describing simple harmonic motion (SHM) for the logarithm of the trait, where is the analog of a spring constant. The “stiffness” of the spring, , is related to the strength of the trait’s effect on fitness. The angular frequency of the motion is determined by the product of with the strength of the maternal effect, , via . In other words,
where and .
The logarithmic population size also oscillates with SHM according to:
so that
Equation (12) shows that, for a certain form of maternal effect, there is a natural relationship between the time derivative of (the logarithm of) the trait value and the logarithm of the population size. By choosing the form of the selection “force” via , we can consider different kinds of “bound motion” of which simple harmonic motion is a fundamental example.
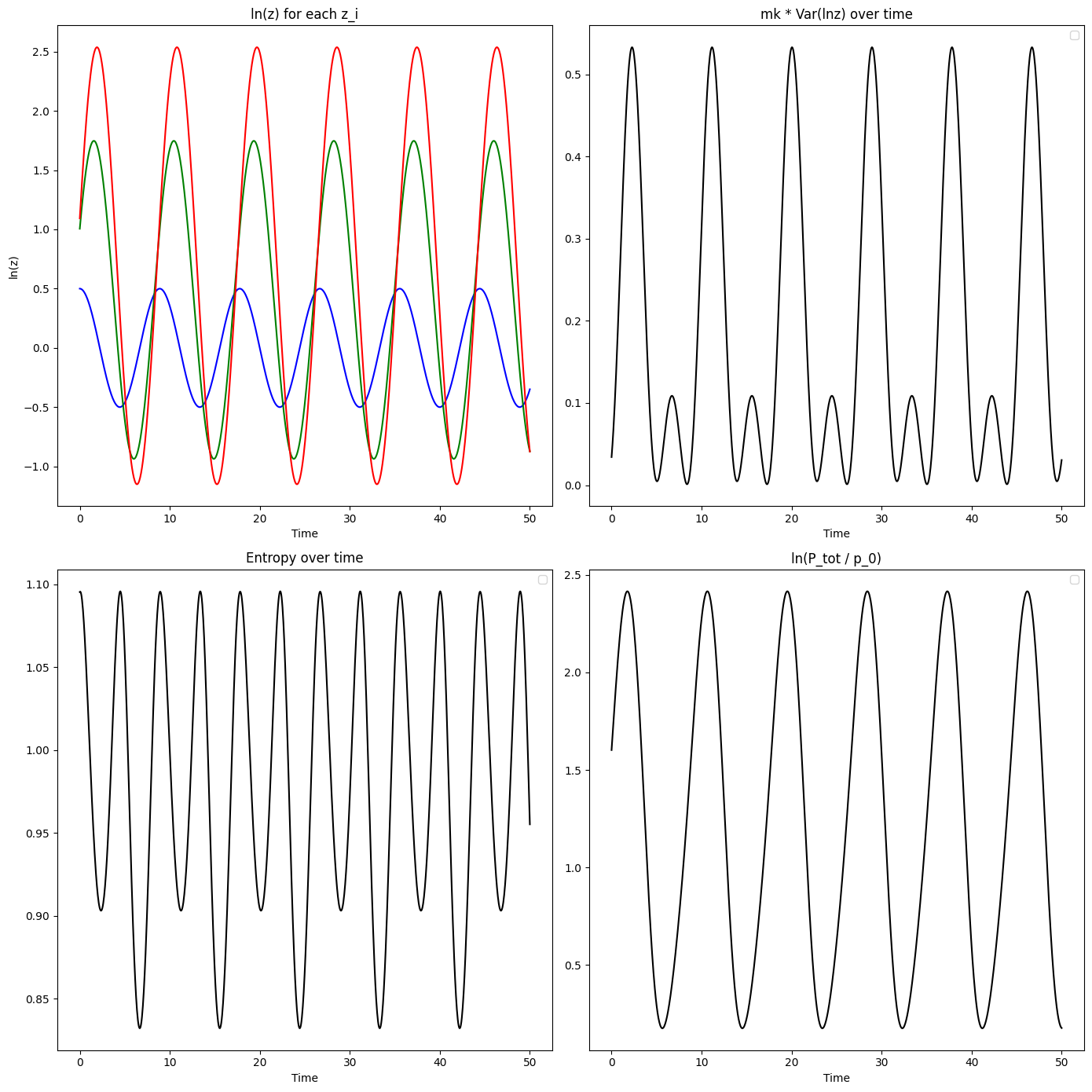
The evolution equations (6,7) and the extension (8) deal with single populations but can be readily extended to multiple subpopulations, which can then be aggregated to shed light on the behavior of a full system. Consider the case in which the subpopulation from among the subpopulations has a value for a trait represented by , a population size , a fitness scaling , and individual maternal effects . Suppose, in addition, that each subpopulation follows SHM as described above. The virial equation (5) applied to is
Since each subpopulation performs SHM in , is bounded, so the time average of will go to zero. From equations (13) and (12), we therefore have:
| (14) | |||||
Note that the standard form of the virial theorem for SHM is an energy equation in and , yielding the relation . On the other hand, (14) is a velocity equation. In the special case where the values , , and are the same between subpopulations and given by , , and , respectively, the above simplifies to
| (15) |
This equation describes the balance between variation in the trait between subpopulations and the entropy in the distribution of subpopulation sizes, when subpopulations are following SHM in the way we have described. When subpopulations have different trait values, selection acts to create a non-uniform distribution of populations sizes. This illustrates the essence of the general case (14), where the Shannon entropy is replaced by a weighted entropy [47].
Notably, the consideration of overall population dynamics resulting from multiple subpopulations exhibiting SHM, leads immediately to a universality observation stemming from the Fourier theorem [43]:
Theorem 5 (Universality of ecological orbits)
Maternal effects driving simple harmonic motion in subpopulations are sufficient to generate any periodic population dynamics that satisfy the Dirichlet conditions.
Evolutionary theory and Newtonian mechanics
The dynamical interpretation of evolutionary theory posits a correspondence between theories of evolution and Newtonian mechanics [46, 23]. In this framework, notions such as selection or mutation in biology are associated to forces in physics [46]. The identical form of equations (4) and (5) can constrain such associations and clarify subsequent analogies. For example, although the standard form of the virial theorem in (2) is an energy equation, equation (5) is a velocity equation, which in biology translates to rates of change of biological quantities. Furthermore, rate of change of a trait or phenotype, i.e., in equation (4) or the finite difference in equation (3), corresponds to the momentum-averaged bulk velocity of a collection of physical objects. The remaining terms in each equation similarly have physical meaning in the context of classical mechanics in equation (5), or biology in equation (3).
| Variable | Biology | Physics | Physical Units |
|---|---|---|---|
| subpopulation | object | 1 | |
| trait / phenotype | position | ||
| evolutionary rate | velocity | ||
| Malthusian fitness | acceleration velocity | ||
| population size | momentum | ||
| relative population size | relative momentum | 1 | |
| population growth rate | force | ||
| population-averaged trait / phenotype | momentum-averaged position | ||
| group evolutionary rate | bulk momentum velocity | ||
| selection rate | extrinsic momentum velocity | ||
| transmission rate | intrinsic momentum velocity |
The momentum-averaged positions and velocities are discrete analogs of momentum-averaged position and momentum velocity in electromagnetism, where they emerge from the virial density in an application of the virial theorem to electromagnetic pulses [11]. Whereas is frequently referred to as the selection term in the Price equation [5], the connection to the virial theorem suggests that it is better described as a selection rate (Table 1). Similarly, the transmission term is more accurately a transmission rate. Most significantly, while the dynamical interpretation typically relies on associating force to natural selection, drift, migration, or mutation [23], the equivalence between the virial theorem and the Price equation, suggests that force is more naturally associated to fitness. The correspondence of force to a rate of change is not surprising, since force is also a rate of change, specifically the rate of change of momentum. This stands more in line with the statistical interpretation of evolutionary theory [54, 53, 29], which, among several critiques of the dynamical interpretation, finds fault with the analogies of biological processes such as mutation with forces in physics, arguing that the physical forces are causal in a way that processes such as selection or mutation are not [54]. However, the analogy of population growth with force can be viewed as consistent with the dynamical interpretation; for example, population growth can directly affect DNA polymorphism patterns [55]. Moreover, the virial theorem in the setting of the ecological simple harmonic oscillator affirms [23] in noting that “natural selection turns out to be more similar to forces such as friction and elastic forces rather than the more canonical gravitation.”
In particular, considerations of the analogies between biology and physics via the virial theorem led us to generalize the work of [18] and to derive the ecological simple harmonic oscillator, which to our knowledge is the first example of such an oscillator that emerges solely from maternal effects and does not require a predator-prey or other more sophisticated model. The extension of (7) to (8) is interesting in its own right, and should be interesting to develop in future work. Moreover, Theorem 5 shows that subpopulations subject to distinct maternal effects can generate arbitrarily complex population dynamics, thereby affirming the main thesis of [18].
Ultimately, analogies between the Price equation and the virial theorem point towards potentially productive directions for exploration in both biology and physics. The statistical framing of the virial theorem in (5) highlights phenomena that may have been overlooked in the physics realm. For example, the first term on the right-hand side of (5), namely , can be understood to quantify the extent of the Yule-Simpson effect [33, 56, 45], which describes a situation where within-group trends can be reversed upon averaging. In biology, the Price equation has the potential to be used more widely as a tool. Although it has been hailed as a unifying framework for researchers [25], one that “can serve as a heuristic principle to formulate and systematize different theories and models in evolutionary biology” [26], the emphasis on its use has been more oriented toward understanding how it generalizes specific equations, rather than applying it for biological discovery. For example, the Price equation can be used to derive the Breeder’s equation [4, 57], Fisher’s fundamental theorem [37, 35], the house of cards approximation for genetic variance at mutation-selection balance [57, 50], and many other formulas and identities in genetics [57, 40]. However, it has been referred to as a tautology and a vacuous statement without application. In [51] the Price equation is described as a theorem that establishes that “If the left-hand side is computed as suggested in [36], and the right-hand side too, then they are equal.” This critique of the Price equation, namely that it does not and cannot serve as a tool, stands in contradiction to evidence from physics, where the mathematically equivalent virial theorem has been understood as a powerful tool since its use to discover dark matter in 1933 [58]. The manifold applications of the virial theorem [28] suggest that there is still much to gain from the application of the Price equation as a tool for biology. In fact, the equivalence we have demonstrated between the Price equation and the virial theorem shows that the description of missing heritability as dark matter [27] may be understood to be more than just an informal analogy between mysteries in genetics and astronomy.
Author contributions
SL studied the virial theorem while participating in the “Introduction to Astrophysics” cluster in the COSMOS summer program held at UC Irvine from July 9, 2023 to August 4, 2023. Specifically, she used the virial theorem to repeat Zwicky’s Coma cluster mass estimates using modern measurements of velocity dispersion and galaxy positions. LP learned of the virial theorem from SL and, in discussing its proof with SL, realized that it must be related to the Price equation. SL and LP explored the applications and implications of the connection. CF identified and developed the connection to ecological orbits and simple harmonic motion via the maternal effect. LP drafted the initial manuscript; both SL and LP edited the first version posted on arXiv [24]. CF, SL, and LP edited the final version and code [12]. The authors are ordered alphabetically.
Acknowledgments
SL thanks Manoj Kaplinghat and Gopolang Mohlabeng who led the “Introduction to Astrophysics” cluster (cluster 4) at the 2023 UC Irvine COSMOS program. LP relied in part on notes about Fisher’s theorem of natural selection from his April 22, 2008 lecture for UC Berkeley course Math 239: Discrete Mathematics for the Life Sciences that were transcribed and edited by Cynthia Vinzant and Caroline Uhler. LP thanks Junhyong Kim for suggesting a possible connection of this work to the ideas in the book Ecological Orbits: How Planets Move and Populations Grow by Lev Ginzburg and Mark Colyvan, a discussion that led to CF’s extension of the work described in the book and the section on simple harmonic motion.
References
- [1] Thomas Alazard and Claude Zuily. Virial theorems and equipartition of energy for water-waves. arXiv preprint arXiv:2304.07872, 2023.
- [2] Esben S Andersen. Population thinking, Price’s equation and the analysis of economic evolution. Evolutionary and Institutional Economics Review, 1:127–148, 2004.
- [3] Jiang-Hui Bian, Shou-Yang Du, Yan Wu, Yi-Fan Cao, Xu-Heng Nie, Hui He, and Zhi-Bing You. Maternal effects and population regulation: maternal density-induced reproduction suppression impairs offspring capacity in response to immediate environment in root voles microtus oeconomus. Journal of Animal Ecology, 84(2):326–336, 2015.
- [4] P Bijma. The price equation as a bridge between animal breeding and evolutionary biology. Philosophical Transactions of the Royal Society B, 375(1797):20190360, 2020.
- [5] Pierrick Bourrat, William Godsoe, Pradeep Pillai, Tarik C Gouhier, Werner Ulrich, Nicholas J Gotelli, and Matthijs van Veelen. What is the price of using the Price equation in ecology? Oikos, page e10024, 2023.
- [6] Rudolf Clausius. XVI. On a mechanical theorem applicable to heat. The London, Edinburgh, and Dublin Philosophical Magazine and Journal of Science, 40(265):122–127, 1870.
- [7] Jacky Cresson, Laurent Nottale, and Thierry Lehner. Stochastic modification of newtonian dynamics and induced potential—application to spiral galaxies and the dark potential. Journal of Mathematical Physics, 62(7), 2021.
- [8] Guillame de L’Hôpital. Analyse Des Infiniment Petits Pour L’Intelligence Des Lignes Courbes. Chez Montalant, Paris, France, 1696.
- [9] Albert Einstein. Die Grundlage der allgemeinen Relativitätstheorie. Annalen der Physik, 49(7), 1922.
- [10] Stephen P Ellner, Monica A Geber, and Nelson G Hairston Jr. Does rapid evolution matter? measuring the rate of contemporary evolution and its impacts on ecological dynamics. Ecology letters, 14(6):603–614, 2011.
- [11] Berthold-Georg Englert. lectures on classical electrodynamics. World Scientific Publishing Company, 2014.
- [12] Catherine Felce, Steinunn Liorsdóttir, and Lior Pachter. Code to reproduce Fig. 1 and Fig. 2.: https://github.com/pachterlab/FLP_2024, 2024. Accessed: 2024-12-30.
- [13] Ronald A Fisher. The genetical theory of natural selection. Clarendon Press, Oxford, Valorium edition, Bennett JH (Editor), 1999, Oxford University Press, Oxford, UK], 1930.
- [14] Ralph H Fowler. Statistical Mechanics. Cambridge University Press, Cambridge, UK, 1929.
- [15] Steven A Frank. The Price equation, Fisher’s fundamental theorem, kin selection, and causal analysis. Evolution, 51(6):1712–1729, 1997.
- [16] Steven A Frank and Montgomery Slatkin. The distribution of allelic effects under mutation and selection. Genetics Research, 55(2):111–117, 1990.
- [17] Vladimir Georgescu and Christian Gérard. On the virial theorem in quantum mechanics. Communications in Mathematical Physics, 208(2):275–281, 1999.
- [18] Lev Ginzburg and Mark Colyvan. Ecological orbits: How planets move and populations grow. Oxford University Press, 2004.
- [19] Lev R. Ginzburg and Dale E. Taneyhill. Population cycles of forest Lepidoptera: A maternal effect hypothesis. Journal of Animal Ecology, 63(1):79–92, 1994.
- [20] Mervin P Hanson. The virial theorem, perfect gases, and the second virial coefficient. Journal of chemical education, 72(4):311, 1995.
- [21] JE Harding. The nutritional basis of the fetal origins of adult disease. International Journal of Epidemiology, 30(1):15–23, 02 2001.
- [22] Oren Harman. The price of altruism: George Price and the search for the origins of kindness. WW Norton & Company, New York, NY, 2011.
- [23] Christopher Hitchcock and Joel D Velasco. Evolutionary and Newtonian forces. Ergo, 1(2):39, 2014.
- [24] Steinunn Liorsdóttir and Lior Pachter. The virial theorem and the price equation. arXiv preprint arXiv:2312.06114 v1, 2023.
- [25] Victor J Luque. One equation to rule them all: a philosophical analysis of the Price equation. Biology & Philosophy, 32(1):97–125, 2017.
- [26] Victor J Luque and Lorenzo Baravalle. The mirror of physics: on how the Price equation can unify evolutionary biology. Synthese, 199(5-6):12439–12462, 2021.
- [27] Teri A Manolio, Francis S Collins, Nancy J Cox, David B Goldstein, Lucia A Hindorff, David J Hunter, Mark I McCarthy, Erin M Ramos, Lon R Cardon, Aravinda Chakravarti, et al. Finding the missing heritability of complex diseases. Nature, 461(7265):747–753, 2009.
- [28] Guilhem Marc and William G McMillan. The virial theorem. Advances in Chemical Physics, pages 209–361, 1985.
- [29] Mohan Matthen and André Ariew. Two ways of thinking about fitness and natural selection. The Journal of Philosophy, 99(2):55–83, 2002.
- [30] David Merritt. The distribution of dark matter in the Coma cluster. The Astrophysical Journal, 313:121–135, 1987.
- [31] Armita Nourmohammad, Torsten Held, and Michael Lässig. Universality and predictability in molecular quantitative genetics. Current opinion in genetics & development, 23(6):684–693, 2013.
- [32] Hasan N Oguz and Andrea Prosperetti. A generalization of the impulse and virial theorems with an application to bubble oscillations. Journal of Fluid Mechanics, 218:143–162, 1990.
- [33] Karl Pearson, Alice Lee, and Leslie Bramley-Moore. VI. Mathematical contributions to the theory of evolution.—VI. Genetic (reproductive) selection: Inheritance of fertility in man, and of fecundity in thoroughbred racehorses. Philosophical Transactions of the Royal Society of London. Series A, Containing Papers of a Mathematical or Physical Character, 6(192):257–330, 1899.
- [34] Paolo Podio-Guidugli. The virial theorem: A pocket primer. Journal of Elasticity, 137(2):219–235, 2019.
- [35] George R Price. Fisher’s ‘fundamental theorem’ made clear. Annals of human genetics, 36(2):129–140, 1972.
- [36] George R Price et al. Selection and covariance. Nature, 227:520–521, 1970.
- [37] David C Queller. Fundamental theorems of evolution. The American Naturalist, 189(4):345–353, 2017.
- [38] Lord Rayleigh. XLII. On the momentum and pressure of gaseous vibrations, and on the connexion with the virial theorem. The London, Edinburgh, and Dublin Philosophical Magazine and Journal of Science, 10(57):364–374, 1905.
- [39] D. Reznick, H. Callahan, and R. Llauredo. Maternal effects on offspring quality in poeciliid fishes. American Zoologist, 36(2):147–156, 2015.
- [40] Sean H Rice. Evolutionary theory: mathematical and conceptual foundations. Sinauer Associates, Sunderland MA, 2004.
- [41] Sean H Rice. A stochastic version of the Price equation reveals the interplay of deterministic and stochastic processes in evolution. BMC evolutionary biology, 8:1–16, 2008.
- [42] Tessa Roseboom, Susanne de Rooij, and Rebecca Painter. The dutch famine and its long-term consequences for adult health. Early Human Development, 82(8):485–491, August 2006. Epub 2006 Jul 28.
- [43] Walter Rudin et al. Principles of mathematical analysis, volume 3. McGraw-hill New York, 1964.
- [44] Steven N Shore. An introduction to astrophysical hydrodynamics. Academic Press, Cambridge, MA, 2012.
- [45] Edward H Simpson. The interpretation of interaction in contingency tables. Journal of the Royal Statistical Society: Series B (Methodological), 13(2):238–241, 1951.
- [46] Elliott Sober. The nature of selection. MIT Press, Cambridge, MA, 1984.
- [47] Yuri Suhov, Izabella Stuhl, Salimeh Yasaei Sekeh, and Mark Kelbert. Basic inequalities for weighted entropies. Aequationes mathematicae, 90:817–848, 2016.
- [48] Lih S The and Simon D M White. The mass of the Coma cluster. Astronomical Journal, 92(6):1248–1253, 1986.
- [49] E Tom Thorne, Ron E Dean, and William G Hepworth. Nutrition during gestation in relation to successful reproduction in elk. The Journal of Wildlife Management, pages 330–335, 1976.
- [50] Michael Turelli. Heritable genetic variation via mutation-selection balance: Lerch’s zeta meets the abdominal bristle. Theoretical population biology, 25(2):138–193, 1984.
- [51] Matthijs Van Veelen, Julián García, Maurice W Sabelis, and Martijn Egas. Group selection and inclusive fitness are not equivalent; the Price equation vs. models and statistics. Journal of theoretical biology, 299:64–80, 2012.
- [52] Günter P Wagner. The measurement theory of fitness. Evolution, 64(5):1358–1376, 2010.
- [53] Denis M Walsh, Tim Lewens, and André Ariew. The trials of life: Natural selection and random drift. Philosophy of Science, 69(3):452–473, 2002.
- [54] Dennis M Walsh. Chasing shadows: natural selection and adaptation. Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, 31(1):135–153, 2000.
- [55] Scott H Williamson, Ryan Hernandez, Adi Fledel-Alon, Lan Zhu, Rasmus Nielsen, and Carlos D Bustamante. Simultaneous inference of selection and population growth from patterns of variation in the human genome. Proceedings of the National Academy of Sciences, 102(22):7882–7887, 2005.
- [56] G Udny Yule. Notes on the theory of association of attributes in statistics. Biometrika, 2(2):121–134, 1903.
- [57] Xu-Sheng Zhang and William G Hill. Change and maintenance of variation in quantitative traits in the context of the price equation. Theoretical population biology, 77(1):14–22, 2010.
- [58] Fritz Zwicky. Die Rotverschiebung von extragalaktischen Nebeln. Helvetica Physica Acta, Vol. 6, p. 110-127, 6:110–127, 1933.